
Chapter 7 Evolution and Speciation
Chapter 7 Outline:
7.1 Charles Darwin and Evolution by Natural Selection
7.2 Processes and Patterns of Evolution
7.4 Speciation – Allopatric and Sympatric Formation of New Species
7.5 Misconceptions about Evolution
7.6 Case Study – Natural Selection in Rock Pocket Mice
Learning Outcomes:
After studying this chapter, each student should be able to:
- 7.1 Describe how the present-day theory of evolution by natural selection was developed
- 7.2 Define adaptation and give examples of patterns of evolution, such as convergent and divergent evolution
- 7.3 Describe various areas of study that support the idea of evolution by natural selection, such as homologous and vestigial structures
- 7.4 Describe how new species can be formed by geographic isolation (allopatric speciation) or in the same area (sympatric speciation)
- 7.5 Discuss misconceptions about the theory of evolution
- 7.6 Explain how changes in coat color among the population of rock pocket mice demonstrates how evolution by natural selection occurs
- 7.7 Summarize the major concepts related to evolution
7.1 Charles Darwin and Evolution by Natural Selection
Evolution by natural selection, an idea first popularized by Charles Darwin (Figure 1), describes a mechanism by which species on our planet have changed over time. This theory states that organisms that are better adapted to their environment are more likely to survive and pass on the genes than those with less favorable traits. This process has is the reason that species have changed and diverged over time. This relatively simple but monumental idea helped explain the existence of such a great variety of species across the globe, and allowed biologists to make sense of the living world.


Figure 1. Two photographs of Charles Darwin, author of On the Origin of Species by Means of Natural Selection (1869) (Credit: Wikimedia Commons).
But in England in the 19th century, most people believed that all known species had been the same since the earth’s beginning. The only commonly read source for information on this topic was the bible, and the Genesis story kept things simple – God created all living things during those first 6 days of the earth! But most scientists and even many educated clergymen of this era knew that the earth itself told a different story. For example, fossils had already been revealed and documented that showed a different menagerie of creatures in times past. So Charles Darwin’s book-length explanation of these changes in species over time, published in 1869 as On the Origin of Species was revolutionary (and heretical) for many, while a welcome explanation of so many biologically puzzling questions for those with a knowledge of science.
The following video gives some deeper background and information on Darwin and the theory of evolution by natural selection:
Bozeman Science (2011, June 21). Natural selection [Video – YouTube]. https://youtu.be/R6La6_kIr9g
Ancient Greeks such as Empedocles thought the world was in a constant cycle of change. In the eighteenth century, ideas about the evolution of animals were reintroduced by the naturalists Georges-Louis Leclerc and Comte de Buffon, who observed that various geographic regions of the earth had different plant and animal populations, even when their environments were similar.
James Hutton, a Scottish naturalist of that day, proposed that the very changes observed by geologists studying the earth in his day may have been occurring over long periods of time, gradually accumulating small changes that resulted in the earth’s current geology. The geologist Charles Lyell (who became a friend to Darwin) applied Hutton’s ideas to the natural world – the notion that the greater age of the Earth gave more time for gradual change in species as well. It was also an accepted idea in the late eighteenth century that certain earlier species had gone extinct, another kind of evolutionary change, and could only be seen today in the form of fossils.
By the mid-nineteenth century, the actual mechanism for evolution was independently conceived of and described by two naturalists: Charles Darwin and Alfred Russel Wallace. Importantly, each naturalist spent time exploring the natural world on expeditions to tropical areas of the earth. From 1831 to 1836, Charles Darwin traveled around the world on H.M.S. Beagle, including stops in South America, Australia, and the southern tip of Africa. Alfred Wallace traveled to Brazil to collect insects in the Amazon rainforest from 1848 to 1852 and to the islands of the Malay Archipelago from 1854 to 1862.
Darwin’s journey, like Wallace’s later journeys to the Malay Archipelago, included stops at several island chains, the last being the Galápagos Islands west of Ecuador. On these islands, Darwin observed species of organisms on different islands that were clearly similar, yet had distinct differences. For example, the ground finches inhabiting the Galápagos Islands comprised several species with unique beak shapes (Figure 2). He observed that these finches closely resembled another finch species on the mainland of South America, yet their beaks were slightly different. Darwin imagined that the island species might be species modified over time from the original mainland species. Upon further study, he realized that the varied beaks of each type of finch helped the birds acquire a specific type of food. For example, seed-eating finches had stronger, thicker beaks for breaking seeds (see Figure 2, #1), and insect-eating finches had spear-like beaks for stabbing their prey (see Figure 2, #4). Darwin postulated that chance changes in the beak shape had allowed certain subpopulations to survive and reproduce better over time, since their beaks were best suited to acquire particular food sources.
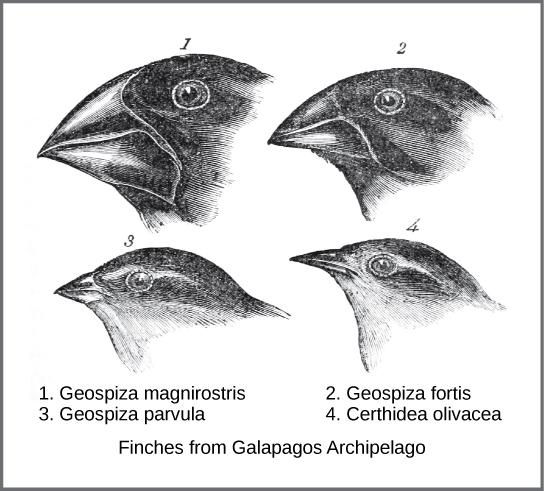
Figure 2. Beak shapes of closely related finches on the Galapagos Islands, as drawn by Charles Darwin. (Credit: Wikimedia Commons).
Both Wallace and Darwin observed similar patterns in other organisms and independently developed the same explanation for how and why such changes could take place. Darwin called this mechanism of evolutionary change over time “natural selection”. Natural selection, sometimes referred to as “survival of the fittest,” states that individuals with favorable traits could survive and reproduce at a higher rate than other individuals. Over time, this led to observable evolutionary changes, with those new, positive traits becoming the norm. With the Galapagos finches, the change in beak shape gave populations a survival advantage, because they could acquire a new food source, which reduced competition for food with the other finches.
In another classic example, a population of giant tortoises found in the Galapagos Archipelago was observed by Darwin to have longer necks than those that lived on other islands with dry lowlands. These tortoises were “selected” because they could reach more leaves and access more food than those with short necks. In times of drought when fewer leaves would be available, those that could reach more leaves had a better chance to eat and survive than those that could not reach the food source. Consequently, long-necked tortoises would be more likely to survive and be reproductively successful than other tortoises, and pass the long-necked trait to their offspring. Over time, only long-necked tortoises would become present in the population.
Natural selection, Darwin argued, was an inevitable outcome of three principles that operated in nature:
Most characteristics of organisms are inherited, or passed from parent to offspring. Although neither Darwin nor Wallace knew how this happened (modern genetics was unknown in their day), the idea that traits were passed from generation to generation was commonly understood.
More offspring are produced than are able to survive, and the resources needed for survival and reproduction are limited. The capacity for reproduction in all organisms outstrips the availability of resources to support their numbers, so better adapted individuals would live while other individuals would die. Both Darwin and Wallace’s understanding of this principle came from reading an essay by the economist Thomas Malthus, who discussed this principle of survival in relation to human populations.
Putting the first two ideas together, Darwin and Wallace stated that variations among members of a species would allow some to survive better than others, passing those favorable traits on to their offspring at a higher rate than less favorable traits. The two biologists reasoned that offspring with inherited characteristics which allow them to best compete for limited resources would survive and have more offspring than those individuals with variations that are less able to compete. Because characteristics are inherited, these traits will be better represented in the next generation. This “survival of the fittest” would lead to changes in populations over generations in a process that Darwin called descent with modification. Ultimately, natural selection leads to greater adaptation of each population to its local environment, a mechanism known as adaptive evolution.
Papers by both Charles Darwin and Alfred Wallace (Figure 3) presenting the idea of natural selection were read together in 1858 before the Linnean Society in London. The following year, Darwin’s book, On the Origin of Species, was published. His book has been described as “a book length argument” for evolution by natural selection, because the book outlines in considerable detail his evidence for evolution by natural selection.

Figure 3. Both (a) Charles Darwin and (b) Alfred Wallace wrote scientific papers on natural selection that were presented together before the Linnean Society in London in 1858. (Credit: National Geographic Education)
Experiments to demonstrate evolution by natural selection can be challenging and time consuming and often involve many years of scientific study. One of the best examples has been demonstrated in the very birds that helped to inspire Darwin’s theory: the Galápagos finches. Peter and Rosemary Grant and their colleagues have studied Galápagos finch populations every year since 1976 and have provided important demonstrations of natural selection. The Grants found changes from one generation to the next in the distribution of beak shapes within populations of medium ground finches on the Galápagos island of Daphne Major. The birds vary in bill shape with some birds having wide deep bills and others having thinner bills. During a period in which rainfall was higher than normal because of an El Niño climate event, the large, hard seeds that large-billed birds ate were reduced in number; however, there was an abundance of the small soft seeds which the small-billed birds ate. Therefore, survival and reproduction were much better in the following years for the small-billed birds. In the years following this El Niño, the Grants measured beak sizes in the population and found that the average bill size was smaller. Since bill size is an inherited trait, parents with smaller bills had more offspring and the size of bills had evolved to be smaller. As conditions improved in 1987 and larger seeds became more available, the trend toward smaller average bill size ceased.
The following video has excellent footage of the Galapagos finches and describes the work of the Grants on this key and formative idea about natural selection:
HHMI BioInteractive (2014, Aug 26). Galapagos Finch Evolution. [Video – YouTube]. https://youtu.be/mcM23M-CCog
Career Connection: Field Biologist
Many people hike, explore caves, scuba dive, or climb mountains for recreation. People often participate in these activities hoping to see wildlife. Experiencing the outdoors can be incredibly enjoyable and invigorating. What if your job was to be outside in the wilderness? Field biologists by definition work outdoors in the “field.” The term field in this case refers to any location outdoors, even under water. A field biologist typically focuses research on a certain species, group of organisms, or a single habitat (Figure 4).

Figure 4. A field biologist tranquilizes a polar bear for study. (credit: Karen Rhode)
One objective of some field biologists is to discover new species that have never been recorded. Not only do such findings expand our understanding of the natural world, but they also lead to important innovations in fields such as medicine and agriculture. Plant and microbial species, in particular, can reveal new medicinal and nutritive understandings. Other organisms can play key roles in ecosystems or be considered rare and in need of protection. When discovered, these important species can be used as evidence for environmental regulations and laws.
7.2. Processes and Patterns of Evolution
Natural selection can only take place if there is variation, or genetic diversity in a population. This means that there are differences among individuals in a population. Importantly, these differences must be genetic, so that it can be passed from generation to the next; otherwise, the selection will not lead to change in the next generation. This is a critical distinction because variation among individuals can also be caused by non-genetic reasons such as an individual being larger because of better nutrition rather than different genes. These non-genetic characteristics are not inherited and therefore do not influence natural selection.
Genetic diversity (variation) comes from two main mechanisms: mutation and sexual reproduction. Mutations, or changes in DNA, are the ultimate source of new genetic variation (new alleles) in any population. The genetic changes caused by mutation can have one of three outcomes on an individual’s fitness, which refers to the organism’s ability to survive and reproduce:
1. A mutation may not affect an individual’s phenotype at all, or, it may affect the individual’s phenotype, but not in a way that affects the individual’s fitness. These are considered neutral mutations.
2. A mutation could affect the phenotype of the organism in a way that reduces its fitness, thus lowering the individual’s ability to survive and reproduce. These are considered detrimental mutations.
3. A mutation may affect the phenotype of the organism in a way that increases its fitness, thus increasing the individual’s ability to survive and reproduce. These are beneficial mutations, which are the basis for natural selection.
Sexual reproduction also leads to genetic diversity. When two parents reproduce, unique combinations of alleles assemble to produce the unique genotypes and associated phenotypes in each of their offspring. In humans, for example, with 46 chromosomes and numerous genes per chromosome, it has been estimated that each child has one of 70 trillion possible genetic combinations! If the newly created combinations of alleles affects the individual’s fitness, then it can influence the evolution of a population. The variety of traits produced by sexually reproducing organisms makes the chance that some of the offspring are favored, surviving and reproducing at higher rates, thereby passing these favorable genetic combinations on to the next generation at higher numbers.
A heritable trait that helps the survival and reproduction of an organism in its present environment is called an adaptation. Scientists describe groups of organisms becoming adapted to their environment when a change in the range of genetic variation occurs over time that increases or maintains the “fit” of the population to its environment. For example, the webbed feet of platypuses are an adaptation for swimming. The snow leopard’s thick fur is an adaptation for living in the cold. The cheetah’s fast speed is an adaptation for catching prey.
Whether or not a trait is favorable depends on the environmental conditions at the time. The same traits are not always selected for because environmental conditions can change. For example, consider a species of plant that grew in a moist climate and did not need to conserve water. Large leaves were selected because they allowed the plant to obtain more energy from the sun than plants with smaller leaves. Large leaves require more water than small leaves, but the moist environment provided favorable conditions to support these large leaves. After thousands of years, however, the climate may have changed, and local precipitation may have been reduced. The direction of natural selection would then shift so that plants with smaller leaves were selected over those with large leaves because those populations were able to conserve water to survive under the new environmental conditions.
The evolution of species over millions of years has resulted in enormous variation in form and function. Sometimes, evolution gives rise to groups of organisms that become different from each other. When two species evolve in diverse directions from a common point, it is called divergent evolution. Such divergent evolution can be seen in the varied forms of reproductive organs of flowering plants. Although their basic anatomy may be similar, these plants have become quite different from each other in terms of reproductive structures as a result of selection in different physical environments and adaptation to different kinds of pollinators (Figure 5).

Figure 5. Flowering plants evolved from a common ancestor. Notice that the (a) Dense Blazing Star Flowering (Liatrus spicata) and the (b) Purple Coneflower (Echinacea purpurea) vary in appearance, yet both share a similar morphology. (Credit a: modification of work by Drew Avery; Credit b: modification of work by Cory Zanker)
In other cases, similar phenotypes evolve independently in distantly related species. For example, flight has evolved in both bats and insects. They both have structures we refer to as wings, which are adaptations to flight, however, the wings of bats and insects have evolved from very different original structures. This phenomenon is called convergent evolution, where similar traits evolve independently in species that do not share a common ancestry. The two species evolved from different ancestors to gain the same function, flying, but did so independently from each other.
Physical changes in organism occur over enormous spans of time and help explain how evolution occurs. Natural selection acts on individual organisms within a varied population, which can then influence and change the characteristics of a species over time. Although natural selection may work in a single generation on an individual, it can take thousands or even millions of years for the genotype of an entire species to evolve. It is over these large time spans that life on earth has changed already and continues to change.
7.3 Evidence of Evolution
The evidence for evolution is compelling and extensive. Looking at every level of organization in living systems, biologists see the signature of past and present evolution. Darwin dedicated a large portion of his book, On the Origin of Species, to identifying patterns in nature that were consistent with evolution, and since Darwin’s time, our understanding has become clearer and broader.
Fossils
Fossils show various remains of living things, and provide solid evidence that organisms from the past are not the same as those found today. Fossils are also useful for following the progression of evolution over time, as one explores layers of rock from different eras. Scientists determine the age of fossils and categorize them from all over the world to determine when the organisms lived relative to each other. The resulting fossil record tells the story of the past and shows the evolution of form over millions of years (Figure 6). For example, scientists have recovered highly detailed records showing the evolution of humans and horses (Figures 6a & 6b). In humans, the skulls have changed dramatically, with cranium sizes generally increasing along with brain size, for example. Ancient horses were not much larger than a medium sized dog, yet the fossil record shows a great increase is size over time leading to today’s horses.

Figure 6. (a) In this display, fossil hominids are arranged from oldest (bottom) to newest (top). As hominids evolved, the shape of the skull changed and enlarged. (b) An artist’s rendition of several extinct species of the genus Equus reveals that these ancient species resembled the modern horse (Equus ferus) but varied in size. (Credit: The Museum of Osteology, Oklahoma City, Oklahoma)
Anatomy
Another type of evidence for evolution is anatomical, such as the presence of homologous structures – structures in different organisms that share the same basic form. For example, as shown below (Figure 7), the bones in the appendages of vertebrates, such as humans, dogs, birds, and whales are all the same. This similarity in appendage bones results from them all sharing a common ancestor, which had these same bones. Over time, evolution led to changes in the shapes and sizes of these bones in different species to best serve various functions, such as grasping, running, flying or swimming, but they have maintained the same basic layout. These synonymous (or homologous) parts would not be the same if these varied organisms had not shared a common history.
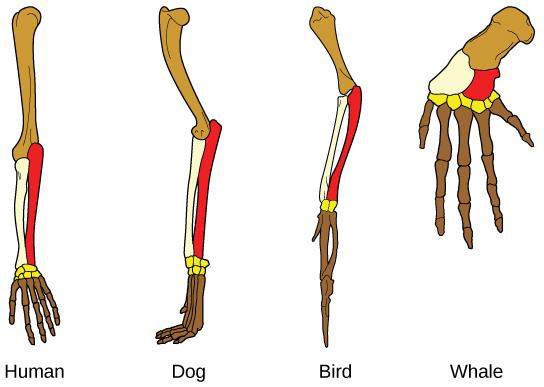
Figure 7. The fact that these dissimilar organisms all have the same bones (though greatly modified) shows that these organisms share a common ancestor. (Credit: Wikimedia Commons).
Some structures exist in organisms that have no apparent current function, called vestigial structures, and appear to be residual (left over) parts from a past ancestors. In other words, these structures only exist because they were functional in past ancestors, and the genes underlying the formation of these structures are still retained. Some easily observed examples of vestigial structures include wings on flightless birds, useless wisdom teeth in humans, and internal hind leg bones in whales and some snakes.
Another area of evidence of evolution related to anatomy is the convergence of form in organisms that share similar environments, called analogous structures. For example, species of unrelated animals, such as the arctic fox and a bird called a ptarmigan, which both live in arctic regions, have been selected for seasonal white phenotypes during winter to blend with the snow and ice (Figures 8a & 8b). These similarities occur not because of common ancestry, but because of similar selection pressures in the same environment, in this case the benefit of not being seen by predators (ptarmigans) or prey (arctic fox). Aloes and euphorbias of the African deserts and agaves and cacti in the deserts of the American continents appear to have similar adaptations to dry conditions, yet are unrelated species. Agaves and aloes have thick and fleshy leaves with sharp thorns that resist both desiccation and herbivory. Cacti and euphorbias have similar adaptations, except that their stems, rather than leaves, have similar adaptations for desert life. Despite evolving in different areas of the world, with different ancestors, similar environmental conditions (hot and dry) have resulted in similar looking adaptations. The similar structures are called analogous structures, while the evolutionary process that lead to analogous structures is called convergent evolution.

Figure 8. The white winter coat of the (a) arctic fox and the (b) ptarmigan’s plumage are adaptations to similar, cold and snowy environments. (Credit: modification of work by Keith Morehouse)
Embryology
Embryology, the study of the development of the anatomy of an organism from embryo to its adult form, also provides evidence of relatedness between now widely divergent groups of organisms. Different vertebrate groups, such as fish, amphibians, reptiles, birds, and mammals, all have similar embryonic structures (e.g., gills and tails) not noted in the adults, showing the common ancestry of these vertebrate groups. Small mutational changes in the embryo can have such magnified consequences in the adult that embryo formation tends to be conserved (not changed over time). As a result, structures that are absent in some vertebrate groups as adults often appear in their embryonic forms and disappear by the time the adult or juvenile form is reached. For example, all vertebrate embryos, including humans, exhibit gill slits and tails at some point in their early development. These disappear in the adults of terrestrial groups but are maintained in adult forms of aquatic groups such as fish and some amphibians. Great ape embryos, including humans, have a tail structure during their development that is lost by the time of birth.
Biogeography
The geographic distribution of organisms (biogeography) across the planet follows patterns that are best explained by evolution in conjunction with the movement of organisms across our planet. These movements have to be understood in terms of a changing positions of land surfaces on our planet over geographic time, as underlying tectonic plates have moved. The study of these evolutionary patterns is called biogeography. For example, groups that evolved before the breakup of the supercontinent Pangaea, when all land surface were joined (about 200 million years ago) are distributed worldwide (Figure 9).
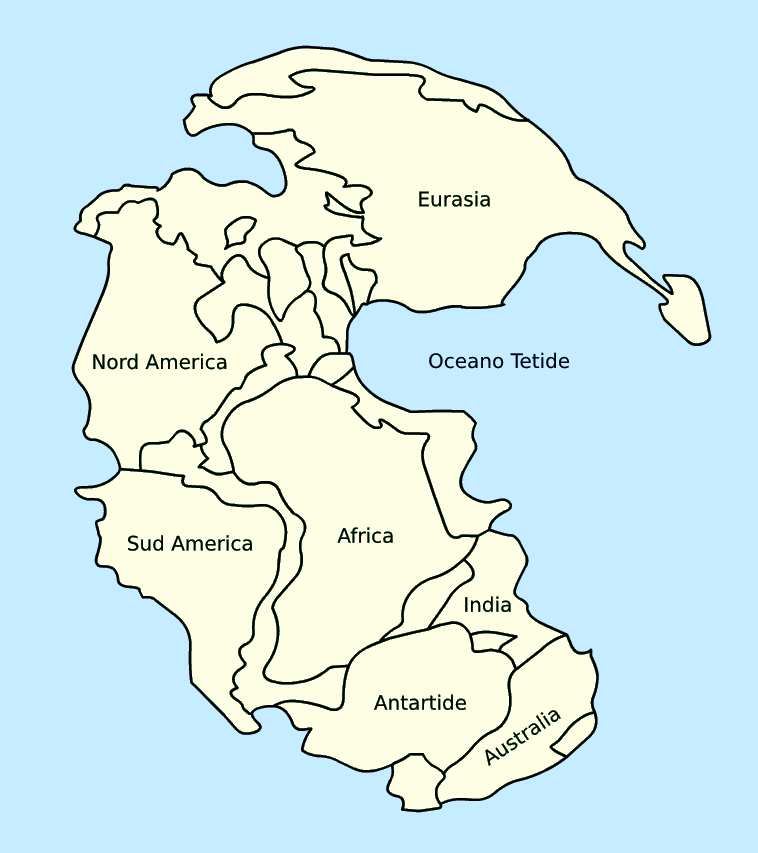
Figure 9. The Supercontinent Pangaea, that existed as a single continent until its break up about 200 million years ago. The breakup was caused by the movement of tectonic plates, which continues today. (Credit: Licensed under the Creative Commons Attribution-Share Alike 3.0 Unported license.)
Groups that evolved after the breakup appear uniquely in regions of the planet, such as the unique flora and fauna of northern continents that formed from the northern supercontinent Laurasia and the southern continents that formed from the supercontinent Gondwana (Figure 10). The presence of members of the plant family Proteaceae in Australia, southern Africa, and South America is best explained by their presence prior to the southern supercontinent Gondwana breaking up.
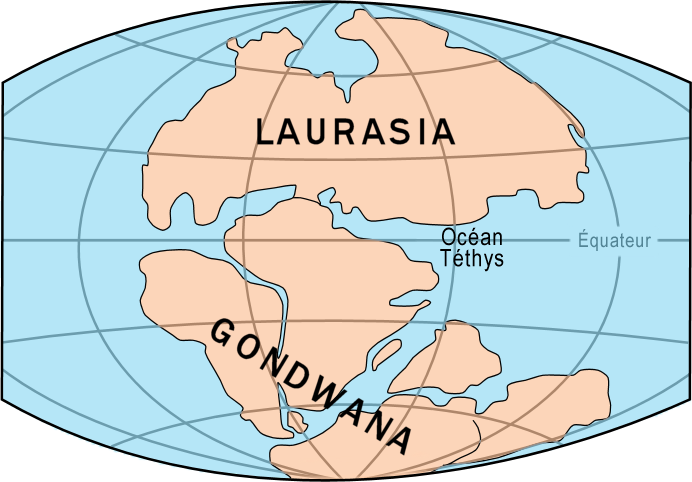
Figure 10. The supercontinents Laurasia and Gondwana, which split off from Pangaea about 200 million years ago, and over the next 150 million years, divided further into the continents we know today. (Credit: This file is licensed under the Creative Commons Attribution-Share Alike 3.0 Unported license.)
The great diversification of marsupials in Australia and the absence of other mammals reflect Australia’s long isolation. Australia has an abundance of endemic species —species found nowhere else—which is typical of islands whose isolation by expanses of water prevents species from migrating. Over time, these species diverge evolutionarily into new species that look very different from their ancestors that may exist on the mainland. The marsupials of Australia, the finches on the Galápagos, and many species on the Hawaiian Islands are all unique to their one point of origin, yet they display distant relationships to ancestral species on mainlands.
Molecular Biology
Like anatomical structures, the structures of the molecules of life reflect descent with modification (evolutionary changes in molecules over time that retain some past features). Evidence of a common ancestor for all of life is reflected in the universality of DNA as the genetic material and in the near universality of the genetic code and the machinery of DNA replication and expression. Fundamental differences between organisms belonging to the various kingdoms of life are reflected in major structural differences, but components of basic structures such as ribosomes and cell membranes show striking similarities. In general, the relatedness of groups of organisms is reflected in the similarity of their DNA sequences—exactly the pattern that would be expected from descent and diversification from a common ancestor.
For example, humans, chimpanzees, and gorillas all descended from a common ancestor, as shown by their genetic similarity. Chimpanzees share 99% of the same DNA as humans, while gorillas are slightly more distant genetically, sharing 98% of human DNA.
DNA sequences have also shed light on some of the mechanisms of evolution. For example, the evolution of new functions for proteins commonly occurs after gene duplication events that allow the free modification of one copy by mutation, while the second copy continues to produce a functional protein.
The following video has excellent photos and illustrations of the evolutionary concepts explored in this section, with an emphasis on the evidence for evolution by natural selection presented by Charles Darwin:
Bozeman Science (2012, March 31). Evidence of Evolution [Video – You Tube] https://youtu.be/cC8k2Sb1oQ8
7.4 Speciation – Allopatric and Sympatric Formation of New Species
The biological definition of species, which works for sexually reproducing organisms, is a group of actually or potentially interbreeding individuals, which produce fertile young. There are exceptions to this rule. Many species are similar enough that hybrid offspring are possible and may often occur in nature, but for the majority of species this rule generally holds. In fact, the presence in nature of hybrids between similar species suggests that they may have descended from a single interbreeding species, and the speciation process may not yet be completed.
Given the extraordinary diversity of life on the planet there must be mechanisms for speciation: the formation of two species from one original species. Darwin envisioned this process as a branching event and diagrammed the process in the only illustration found in On the Origin of Species (Figure 11a). Compare this illustration to the diagram of elephant evolution (Figure 11b), which shows that as one species changes over time, it branches to form more than one new species, repeatedly, as long as the population survives or until the organism becomes extinct.

Figure 11. The only illustration in Darwin’s On the Origin of Species is (a) a diagram showing speciation events leading to biological diversity. The diagram shows similarities to phylogenetic charts that are drawn today to illustrate the relationships of species. (b) Modern elephants evolved from the Palaeomastodon, a species that lived in Egypt 35–50 million years ago. (Credit: OER Commons.org)
For speciation to occur, two new populations must be formed from one original population and they must evolve in such a way that it becomes impossible for individuals from the two new populations to interbreed. Biologists have proposed mechanisms by which this could occur that fall into two broad categories:
(1) Allopatric speciation (allo- = “other”; -patric = “homeland”) involves geographic separation of populations from a parent species and subsequent evolution of two separate species.
(2) Sympatric speciation (sym- = “same”; -patric = “homeland”) involves speciation occurring within a parent species remaining in one location.
Biologists think of speciation events as the splitting of one ancestral species into two descendant species. There is no reason why there might not be more than two species formed at one time except that it is less likely and multiple events can be conceptualized as single splits occurring close in time.
Allopatric Speciation
A geographically continuous population has a gene pool that is relatively homogeneous. Gene flow, the movement of alleles across the range of the species, is relatively free because individuals can move and then mate with individuals in their new location. Thus, the frequency of an allele at one end of a distribution will be similar to the frequency of the allele at the other end. When populations become geographically discontinuous, that free-flow of alleles is prevented. When that separation lasts for a period of time, the two populations are able to evolve along different trajectories. Thus, their allele frequencies at numerous genetic loci gradually become more and more different as new alleles independently arise by mutation in each population. Typically, environmental conditions, such as climate, resources, predators, and competitors for the two populations will differ causing natural selection to favor divergent adaptations in each group.
Isolation of populations leading to allopatric speciation can occur in a variety of ways: a river forming a new branch, erosion forming a new dividing valley or canyon, a group of organisms traveling to a new location without the ability to return (such as by floating on vegetation across an expanse of water), or seeds floating over the ocean to an island. The nature of the geographic separation necessary to isolate populations depends entirely on the biology of the organism and its potential for dispersal. If two flying insect populations took up residence in separate nearby valleys, chances are, individuals from each population would fly back and forth continuing gene flow. However, if two rodent populations became divided by the formation of a new lake, continued gene flow would be unlikely; therefore, speciation would be more likely.
Biologists group allopatric processes into two categories: dispersal and vicariance. Dispersal is when a few members of a species move to a new geographical area, and vicariance is when a natural situation arises to physically divide organisms, such as the formation of a canyon or the rising of a mountain range.
Scientists have documented numerous cases of allopatric speciation taking place. For example, along the west coast of the United States, two separate subspecies of spotted owls exist. The northern spotted owl has genetic and phenotypic differences from its close relative: the Mexican spotted owl, which lives in the south (Figure 12).
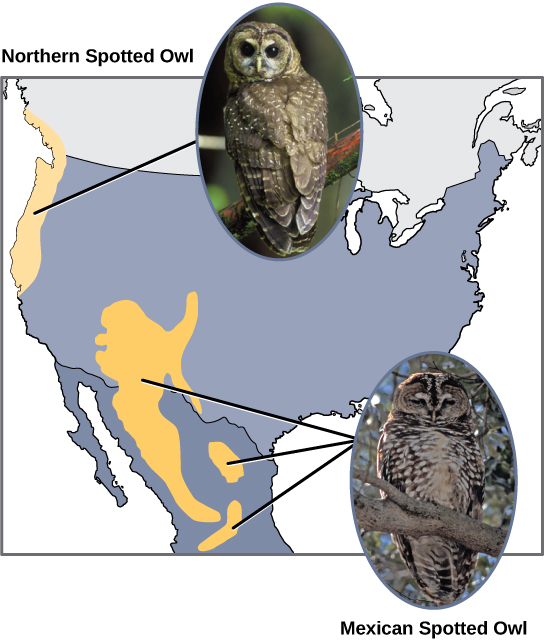
Figure 12. The northern spotted owl and the Mexican spotted owl inhabit geographically separate locations with different climates and ecosystems. The owl is an example of allopatric speciation. (Credit: OER Commons.org.; credit “northern spotted owl”: modification of work by John and Karen Hollingsworth; credit “Mexican spotted owl”: modification of work by Bill Radke)
Additionally, scientists have found that the further the distance between two groups that once were the same species, the more likely it is that speciation will occur. This seems logical because as the distance increases, the various environmental factors would likely have less in common than locations in close proximity. Consider the two owls: in the north, the climate is cooler than in the south; the types of organisms in each ecosystem differ, as do their behaviors and habits; also, the hunting habits and prey choices of the southern owls vary from the northern owls. These variances can lead to evolved differences in the owls, and speciation likely will occur.
Adaptive Radiation – allows for allopatric speciation throughout an area
In some cases, a population of one species disperses throughout an area, and subsets of the population find distinct niches or isolated habitats, eventually forming new species. Over time, the varied demands of their new lifestyles and separated locations may lead to multiple speciation events originating from a single species. This is called adaptive radiation because many adaptations evolve from a single point of origin; thus, causing the species to radiate into several new ones. Island archipelagos like the Hawaiian Islands provide an ideal context for adaptive radiation events because water surrounds each island which leads to geographical isolation for many organisms. The Hawaiian honeycreeper illustrates one example of adaptive radiation. From a single species, called the founder species, numerous species have evolved, including the six shown in Figure 13.
Notice the differences in the species’ beaks in Figure 13. Evolution in response to natural selection based on specific food sources in each new habitat led to evolution of a different beak suited to the specific food source. The seed-eating bird has a thicker, stronger beak which is suited to break hard nuts. The nectar-eating birds have long beaks to dip into flowers to reach the nectar. The insect-eating birds have beaks like swords, appropriate for stabbing and impaling insects. Darwin’s finches are another example of adaptive radiation in an archipelago.
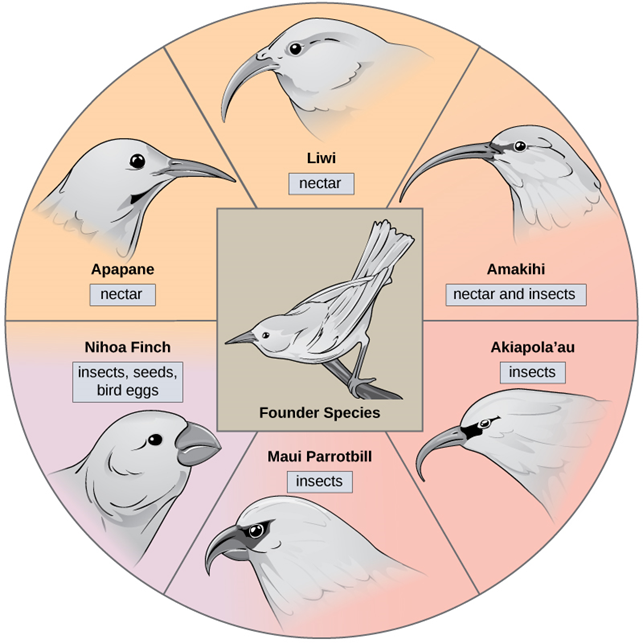
Figure 13. The honeycreeper birds illustrate adaptive radiation. From one original species of bird, multiple others evolved, each with its own distinctive characteristics. (Credit: OER Commons.org)
Sympatric Speciation
Can divergence occur if no physical barriers are in place to separate individuals who continue to live and reproduce in the same habitat? The answer is yes. The process of speciation within the same area is called sympatric speciation; the prefix “sym” means same, so “sympatric” means “same homeland” in contrast to “allopatric” meaning “other homeland.” A number of mechanisms for sympatric speciation have been proposed and studied.
Behavioral Isolation
Behavioral isolation occurs when the presence or absence of a specific behavior prevents reproduction from taking place. For example, male fireflies use specific light patterns to attract females. Various species of fireflies display their lights differently. If a male of one species tried to attract the female of another, she would not recognize the light pattern and would not mate with the male.
Sometimes animals living in the same geographic area may be separated by behavioral differences. One example of this can be found in two mouse species inhabiting the mountains of Appalachia. Peromyscus maniculatus is the white-footed mouse that is largely arboreal (tree dwelling). Peromyscus leucopus, called a deer mouse, lives primarily on the ground. It has been hypothesized that these two species formed from a shared ancestor by this separation in preferred niches, so that they no longer were in contact to mate with each other. Over time they changed enough to no longer be able to mate and produce fertile offspring, so that the two species became distinct.
Chromosomal Errors
Aneuploidy: Aneuploidy is a serious chromosomal error during cell division that can lead to sympatric speciation. In a normal cell division event chromosomes replicate, pair up, and then separate so that each new cell has the same number of chromosomes. However, sometimes the pairs separate and the end cell product has too many or too few individual chromosomes in a condition called aneuploidy, potentially forming a different species (Figure 14).
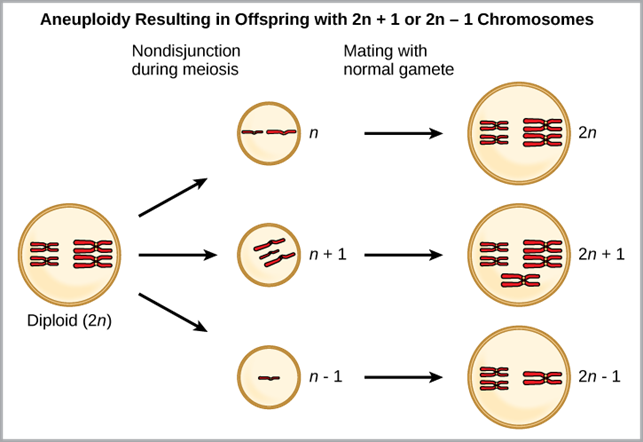
Figure 14. Aneuploidy results when the gametes have too many or too few chromosomes due to nondisjunction during meiosis. In the example shown here, the resulting offspring will have 2n+1 or 2n-1 chromosomes. (Credit: OER Commons.org)
Polyploidy: Polyploidy is a condition in which a cell or organism has an extra set, or sets, of chromosomes, that can also lead to sympatric speciation. Scientists have identified two main types of polyploidy that can lead to reproductive isolation of an individual in the polyploidy state. Reproductive isolation is the inability to interbreed. In some cases, a polyploid individual will have two or more complete sets of chromosomes from its own species in a condition called autopolyploidy. The prefix “auto-” means “self,” so the term means multiple chromosomes from one’s own species. Polyploidy results from an error in meiosis in which all of the chromosomes move into one cell instead of separating.
Autopolyploidy: Autopolyploidy is a condition that results when mitosis is not followed by cell division (cytokinesis) (Figure 15). For example, if a plant species with 2n = 6 produces autopolyploid gametes that are also diploid (2n = 6, when they should be n = 3), the gametes now have twice as many chromosomes as they should have. These new gametes will be incompatible with the normal gametes produced by this plant species. However, they could either self-pollinate or reproduce with other autopolyploid plants with gametes having the same diploid number. In this way, sympatric speciation can occur quickly by forming offspring with 4n called a tetraploid. These individuals would immediately be able to reproduce only with those of this new kind and not those of the ancestral species.
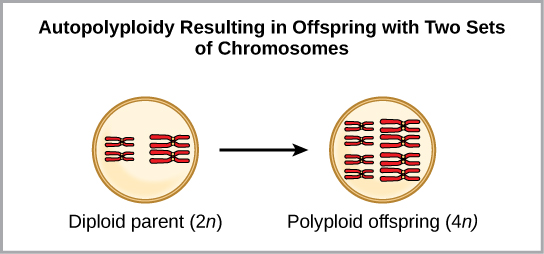
Figure 15. Autopolyploidy resulting from mitosis (which produces two sets of chromosomes in readiness for cell division), that is not actually followed by cell division (cytokinesis). (Credit: OER Commons.org)
Alloploidy: Alloploidy is a condition that results when two different species reproduce to produce viable offspring. In the example shown (Figure 16), a normal gamete from one species fuses with a polyploidy gamete from another. Two matings are necessary to produce viable offspring.
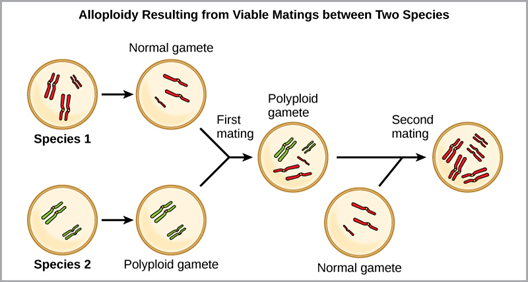
Figure 16. Alloploidy resulting in a viable offspring, although parents are from different species. This is more common in plants than in animals. (Credit: OER Commons.org)
The cultivated forms of wheat, cotton, and tobacco plants are all allopolyploids. Although polyploidy could conceivably occur in animals, it rarely does (animals with any of the types of chromosomal aberrations described here are unlikely to survive and produce normal offspring). Alloploidy takes place most commonly in plants. Scientists have discovered more than half of all plant species studied relate back to a species evolved through polyploidy. With such a high rate of polyploidy in plants, some scientists hypothesize that this mechanism takes place more as an adaptation than as an error.
Reproductive Isolation
Given enough time, reproductive isolation may occur as genetic and phenotypic divergence between populations affects characteristics that influence reproduction. If individuals of the two populations were to be brought together at that point, mating would be less likely, and even if mating occurred, offspring would be non-viable or infertile. At that point in reproductive isolation two new species have formed! Many types of diverging characters may affect the reproductive isolation, the ability to interbreed, of the two populations.
Reproductive isolation can take place in a variety of ways. Scientists organize them into two groups: prezygotic barriers and postzygotic barriers. Recall that a zygote is a fertilized egg: the first cell of the development of an organism that reproduces sexually. Therefore, a prezygotic barrier is a mechanism that blocks reproduction from taking place; this includes barriers that prevent fertilization when organisms attempt reproduction. A postzygotic barrier occurs after zygote formation; this includes young that do not survive the embryonic stages or those that are born sterile.
Prezygotic Barriers
Temporal Isolation
Some types of prezygotic barriers prevent reproduction entirely. Many organisms only reproduce at certain times of the year, often just annually. Differences in breeding schedules, called temporal isolation, can act as a form of reproductive isolation. For example, two species of frogs inhabit the same area, but one reproduces from January to March, whereas the other reproduces from March to May (Figure 17).

Figure 17. These two related frog species exhibit temporal reproductive isolation. (a) Rana aurora breeds earlier in the year than (b) Rana boylii. (Credit: OER commons.org; a: modification of work by Mark R. Jennings, USFWS; b: modification of work by Alessandro Catenazzi)
Habitat Isolation or Division
In some cases, populations of a species move or are moved to a new habitat and take up residence in a part of the habitat that no longer overlaps with the other populations of the same species. This situation is called habitat isolation. Reproduction with the parent species ceases, and a new group exists that is now reproductively and genetically independent. For example, a cricket population that was divided after a flood could no longer interact with each other. Over time, the forces of natural selection, mutation, and genetic drift resulted in the divergence of the two groups (Figure 18).
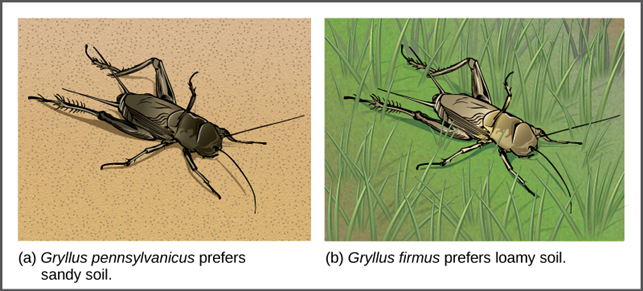
Figure 18. Speciation can occur when two populations occupy different habitats. The habitats need not be far apart. The cricket (a) Gryllus pennsylvanicus prefers sandy soil, and the cricket (b) Gryllus firmus prefers loamy soil. The two species can live in close proximity, but because of their different soil preferences, they became genetically isolated. (Credit: OER Commons.org)
Another example of habitat division and subsequent speciation has been noted for species of fish living in a lake. As the population grew in this confined area, competition for food also grew. Under pressure to find food, a group of these fish may have had the genetic flexibility to discover and feed off a distinct resource that was unused by the other fish, at a different depth in the lake. Over time, those feeding on the second food source interacted more with each other than the original population, preferentially breeding together. Offspring of these fish would likely behave as their parents: feeding, living, and breeding in the same area and keeping separate from the original population. If this group of fish remained separate from the first population, eventually sympatric speciation might occur as more genetic differences accumulated between them.
One such place that this has occurred is Lake Victoria in Africa, famous for its sympatric speciation of cichlid fish. Researchers have found hundreds of sympatric speciation events in these fish, which have not only happened in greater numbers, but also over a shorter period of time.
The two fish in the following diagram show speciation among a cichlid fish population in volcanic crater lakes in Nicaragua (Figure 19). In this locale, two types of cichlids live in the same geographic location but have come to have different morphologies that allow them to eat various food sources.

Figure 19. Cichlid fish from Lake Apoyeque, Nicaragua, show evidence of sympatric speciation. Lake Apoyeque, a crater lake, is 1800 years old, but genetic evidence indicates that the lake was populated only 100 years ago by a single population of cichlid fish. Nevertheless, two populations with distinct morphologies and diets now exist in the lake, and scientists believe these populations may be in an early stage of speciation. (Credit: OER Commons.org)
Gametic Barriers
Other prezygotic barriers work when differences in their gamete cells (eggs and sperm) prevent fertilization from taking place; this is called a gametic barrier. Similarly, in some cases closely related organisms try to mate, but their reproductive structures simply do not fit together. For example, damselfly males of different species have differently shaped reproductive organs. If the male of one species tries to mate with the female of another species, their body parts simply do not fit together (Figure 20).

Figure 20. The shape of the male reproductive organ (penis) varies among male damselfly species and is only compatible with the female of that species. Reproductive organ incompatibility keeps the species reproductively isolated. (Credit: OER Commons.org)
In plants, certain structures aimed to attract one type of pollinator simultaneously prevent a different pollinator from accessing the pollen. The tunnel through which an animal must access nectar can vary widely in length and diameter, which prevents the plant from being cross-pollinated with a different species (Figure 21).
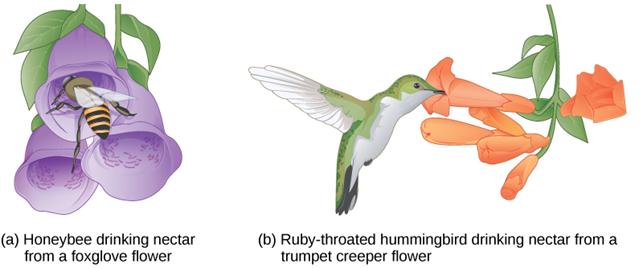
Figure 21. Some flowers have evolved to attract certain pollinators. The (a) wide foxglove flower is adapted for pollination by bees, while the (b) long, tube-shaped trumpet creeper flower is adapted for pollination by humming birds. (Credit: OER Commons.org)
Postzygotic Barriers
Postzygotic barriers can prevent reproduction when fertilization takes place and a zygote forms. Hybrid individuals in many cases cannot form normally in the uterus and simply do not survive past the embryonic stages. This is called hybrid inviability because the hybrid organisms simply are not viable.
In another postzygotic situation, reproduction leads to the birth and growth of hybrid young that are sterile and unable to reproduce offspring of their own, called hybrid sterility. A horse and a donkey can mate and produce a mule, but a mule is sterile (Figure 22).
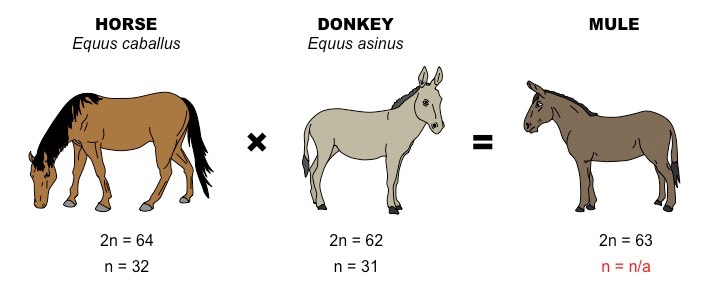
Figure 22. A horse and a donkey can breed to produce a mule as an offspring. The mule, however, is sterile, making the horse and donkey two reproductively isolated species. (Credit: OER Commons.org)
7.5. Misconceptions about Evolution
Although the theory of evolution generated some controversy when it was first proposed, it was almost universally accepted by biologists within 20 years after the publication of On the Origin of Species. Nevertheless, the theory of evolution is a difficult concept and misconceptions about how it works abound.
Evolution misconception 1: There are “missing links” in the fossil record.
When Darwin’s theories were first proposed, there were many “missing links” (absence of transitional forms) in the fossil record of his time. Since then, however, a rich history of fossil organisms have been found, such as many species of therapsids (Figure 23), having characteristics midway between mammals and the reptiles they evolved from. Most “missing links” are no longer missing!

Figure 23. Artists rendering of a therapsid, a fossil organism that evolved from a reptile and later evolved into a mammal, one of the many “missing links” that have been found since the time of Charles Darwin. (Credit: Creative Commons Attribution-Share Alike 4.0 International license.
Evolution misconception 2: Evolution is just a “theory”.
Critics of the theory of evolution dismiss its importance by purposefully confounding the everyday usage of the word “theory” with the way scientists use the word. In science, a theory is understood to be a body of thoroughly tested and verified explanations for a set of observations of the natural world. Scientists have a cell theory, atomic theory, and the theory of relativity, each of which describes understood facts about the world, based on many years of experimentation. In the same way, the theory of evolution describes the living world as we know it. As such, a theory in science has survived significant efforts to discredit it by scientists. In contrast, a “theory” in common vernacular is a word meaning a guess or suggested explanation; this meaning is more akin to the scientific concept of “hypothesis.” When critics of evolution say evolution is “just a theory,” they are implying that there is little evidence supporting it and that it is still in the process of being rigorously tested. This is a mischaracterization, since strong support of this theory comes from so many different realms of scientific inquiry.
Evolution misconception 3: Individuals evolve.
Evolution is the change in genetic composition of a population over time, specifically over generations, resulting from differential reproduction of individuals with certain alleles. Individuals do change over their lifetime, but this is called development and involves changes programmed by the set of genes the individual acquired at birth in coordination with the individual’s environment. When thinking about the evolution of a characteristic, it is probably best to think about the change of the average value of the characteristic in the population over time. For example, when we say that natural selection leads to bill-size change in medium-ground finches in the Galápagos, this does not mean that the individual bills on each currently living bird are changing. Instead, this means that If one measures the average bill size among all individuals in the entire population at one time and then measures the average bill size in the population many years later, this average value may be different as a result of evolution. This change may be brought about by selective pressure; that is birds in the population with more functional bills survive and reproduce at a higher rate than the others, causing the shift in average population wide bill size.
Evolution misconception 4: Evolution explains the origin of life.
It is a common misunderstanding that evolution includes a full explanation of life’s origins. Conversely, some of the theory’s critics believe that it cannot explain the origin of life, so must be invalid. The theory of evolution does not directly explain the origin of life. Instead, this theory explains how populations change over time and how living things have diversified since the earliest species. The mechanisms of the origin of life on Earth are a particularly difficult problem because it occurred a very long time ago, and we have fossils of the earliest cells, but not the process that brought them into existence.
However, once a mechanism of inheritance was in place in the form of a molecule like DNA either within a cell or some form of pre-cell, these entities would be subject to the principle of natural selection. More effective survivors and reproducers would increase in frequency at the expense of less efficient organisms. So while evolution does not explain how that first cell appeared, the theory can be applied to the processes operating once pre-living entities acquired certain properties.
Many hypotheses do exist regarding the appearance of the first cells, that are based on what we know about the energy present at that time (e.g., heat of the earth, light, unfiltered ultraviolet rays from the sun, etc.), as well as the gases present in the early earth’s atmosphere (e.g. methane, ammonia, water vapor, hydrogen, etc.). This combination of energy sources and gases may have been what was necessary to allow early cells to form, since many millions of years passed with these resources and energy forms available, and the earth is a very large laboratory! As you can imagine, research in this area is ongoing.
Evolution misconception 5: Organisms evolve on purpose.
Statements such as “organisms evolve in response to a change in an environment” are quite common, but such statements can lead to two types of misunderstandings. First, the statement must not be understood to mean that individual organisms evolve. The statement should more accurately be written as “a population evolves in response to a changing environment.” However, a second misunderstanding may arise by interpreting the statement to mean that the evolution is somehow intentional. Evolution by natural selection is actually a random event. Random genetic mutations lead to changes in an organism’s phenotype. If the phenotypic change enhances the individual’s ability to survive and reproduce in the current environment, then that individual’s genes are more likely to be passed onto the next generation. If the random mutation does not provide a reproductive benefit, then the phenotype will not be more frequent in the next generation. The environment also changes over time, so random mutations that are beneficial in an environment at a given time may no longer be beneficial in a future environment that has changed. Environments also differ between geographic areas, so genetic mutations that are beneficial in the environment of one location may not be beneficial in the environment of a different location.
To reiterate, it is important to understand that the variation that natural selection works on is already in a population and does not arise in response to an environmental change. For example, applying antibiotics to a population of bacteria will, over time, select a population of bacteria that are resistant to antibiotics. The resistance, which is caused by a gene, did not arise because of the application of the antibiotic. The gene for resistance was already present in the gene pool of the bacteria (perhaps from an earlier mutation), but likely existed at a low frequency. The antibiotic, which kills the bacterial cells without the resistance gene, strongly selects for individuals that are resistant, since these would be the only ones that survived the application of the antibiotics and were able to reproduce. Experiments have demonstrated that mutations for antibiotic resistance do not arise as a result of the antibiotic.
In a larger sense, evolution is not “goal” directed. Species do not become “better” over time; they simply track their changing environment with adaptations that maximize their survival and reproduction in a particular environment at a particular time. Evolution has no goal of making faster, bigger, more complex, or even smarter species, despite the commonness of this kind of language in popular discourse. What characteristics evolve in a species are a function of the genetic variation present in a population and the current state of that population’s environment, both of which are constantly changing in a non-directional way. A trait that is fit in one environment at one time may well be fatal at some point in the future. This holds equally well for a species of insect as it does for the human species.
7.6. Case Study – Evolution by Natural Selection of Rock Pocket Mice
This resource is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International license. No rights are granted to use HHMI’s or BioInteractive’s names or logos independent from this Resource or in any derivative works.
Evolution by natural selection is briefly described as “survival of the fittest”. This is a key evolutionary process whereby individuals of a population with more favorable traits survive and reproduce at a higher rate than individuals without those traits. This process of change in a population can take thousands or even millions of years.
An interesting example of natural selection has been documented by scientists studying the rock pocket mice living in the deserts of New Mexico and Mexico. These mice show a number of coat colors over their range, but two common colors are a lighter tawny brown and a dark black. Predators of this mouse, such as owls, locate the mice mostly by sight, so the mice try to avoid predation by hiding among the rocks.
This video shows an example of how natural selection has occurred among a population of rock pocket mice in the desert of New Mexico:
HHMI BioInteractive (2014, Aug 26). Natural selection and the rock pocket mouse. [Video – YouTube]. https://youtu.be/sjeSEngKGrg
Certain areas of the desert have light colored desert rocks and soil, while other areas have much darker basaltic rock that came from lava flows. In some areas, there are even “islands” of dark rock and soil within larger lighter colored regions.
Scientists have studied populations of rock pocket mice throughout this desert habitat. What they have found is that the light colored (tawny) mice are almost exclusively living in lighter rock areas. The dark colored mice, in contrast, are almost always found on the dark lava rocks and soil. This differentiation occurred even on the dark basaltic islands that were surrounded by lighter rock.
This is an excellent example of how evolution by natural selection works. In the lighter areas, the dark mice stood out and were much more likely to be eaten by predators. The tawny mice, in contrast, were difficult to see against this lighter background and survived and reproduced at a higher rate than the darker mice. Similarly, in the darker areas, the blacker mice were more successful, surviving and reproducing at a higher rate, while the lighter mice stood out and fell victim to predators more often.
These observations have led to extensive genetic studies as well. The underlying genes for producing coat color have been identified and linked to the two major coat colors. Interestingly, the gene producing the dark coat is not the same gene in Mexico as it is in New Mexico, indicating that the two populations experienced different mutations that resulted in the same phenotype, giving both populations the same selective advantage on the dark colored substrate. Although these “melanic” genes for black coats were probably rare before the lava flows, in both locations these rare genes were selected for because of predation, allowing the mice in these areas to survive and reproduce better, because the black coated mice matched the dark background, and were harder for predators to locate and eat (Figure 24).

Figure 24. Rock pocket mice with adaptive coloration on the desert floor they live on. (Credit: National Park Service)
Many traits that have undergone natural selection are more subtle than coat color, but natural selection works in a similar manner, allowing those individuals with better traits (e.g., higher intelligence, better eyesight, more thorns, etc.) to survive and reproduce at higher rates, while those individuals with lesser traits eventually die out in the population.
In today’s world with strong human influences, traits such as smaller tusks on elephants and quieter rattles on rattlesnakes have been selected for. This is because elephants with larger tusks are killed at a higher rate for their valuable ivory, allowing more smaller tusked elephants to survive and reproduce. Rattlesnakes that reveal their presence to humans with a loud rattle are killed more often than the quiet ones, making the quieter rattlesnakes more successful over time!
In the area of health care, antibiotic resistant bacteria have evolved from populations that were mostly susceptible to antibiotics when the antibiotic was first introduced. Only the resistant strains of bacteria were able to survive and reproduce, while the more susceptible strains died out.
7.7. Summary
Evolution is the process of adaptation that occurs because mutations and variable traits are present in all populations. This natural variation allows organisms with more desirable characteristics to survive and reproduce at a higher rate than others, passing on these desirable traits to the next generation at a higher rate than less desirable characteristics are passed on. Over time, populations may evolve that have traits that are beneficial to the overall survival of organisms in the population. For living organisms to adapt and change in response to environmental pressures, genetic variation must be present, which has its origin in naturally occurring genetic mutations. With genetic variation, individuals have differences in form and function that allow some to survive certain conditions better than others. These organisms pass their favorable traits to their offspring. Our earth is always changing, and with that change comes changes in the environments in which organisms live. Over time, what was once a desirable, advantageous trait may become an undesirable trait, or visa versa. Organisms on our changing earth may either evolve, if positive traits are available, or go extinct, if the changes in the earth are not matched by the traits available in a population. Evolution is described as convergent when similar traits evolve in multiple, often unrelated species. Divergent evolution occurs when different traits are shown in multiple species that all came from a common ancestor. Some of the more common pieces of evidence for evolution can be noted by an exploration of the universal DNA code, by the fossil record, and by the existence of homologous structures and vestigial structures.
Attribution:
Content in this chapter includes original work created by Lauren Roberts and Paul Bosch as well as from the following sources, with some modifications:
OER Commons – Biology – Evolutionary Processes https://www.oercommons.org/courseware/lesson/15029/overview

{kind=link}